Acaulospora mellea
(reference accession BR983A)
Whole Spores

 COLOR: Pale orange-brown (0-20-40-0) to dark orange-brown (0-40-100-10), most orange-brown (0-40-80-0).
COLOR: Pale orange-brown (0-20-40-0) to dark orange-brown (0-40-100-10), most orange-brown (0-40-80-0).
SHAPE: Mostly globose, subglobose, occasionally irregular.
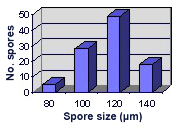
SIZE DISTRIBUTION: 90-140 µm, mean = 120 µm (n = 142)
Subcellular Structure of Spores
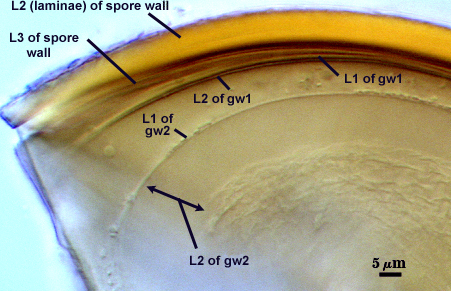
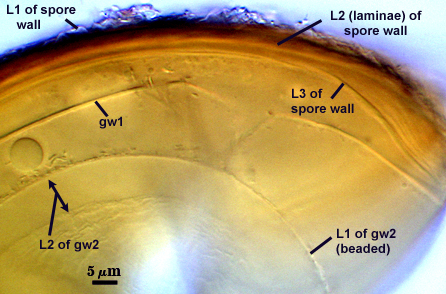
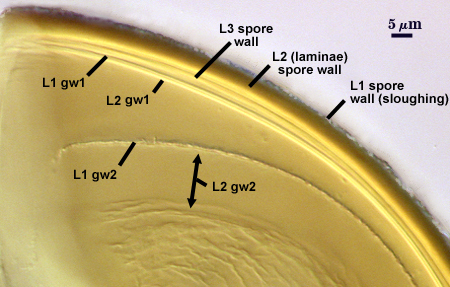
SPORE WALL: Three layers (L1, L2, and L3), the outer continous with the wall of the neck of the parent sporiferous saccule and the latter being synthesized with development of the spore.
L1: Hyaline, thin (< 0.5 µm), and so flexible that it often produces extensive wrinkles in PVLG. Sloughs on many spores, particularly those collected from the field or stored pot cultures. When this layer does not slough, it produces numerous folds on the spore surface and appears “rugose”. This layer has been defined as a “membranous wall”, but it clearly has a different origin and relationship with neighboring structures than inner flexible walls.
L2: A layer consisting of very fine and adherent sublayers (or laminae), pale orange-yellow (0-10-60-0), 3.6-5.1 µm thick (mean = 4.3 µm). Surface is smooth if the outer layer has sloughed, which usually occurs (especially after sucrose-density centrifugation and washing). At maturity, the pore between spore and saccule neck is closed by continuous sublayers of this layer encloses the spore contents in an “endospore”.
L3: Pale yellow-brown (0-5-40-0), thin (1- 4 µm) and somewhat flexible when it separates from L2 of the spore wall. This layer also appears to consist of sublayers (laminae) which can separate more readily from each other than those of L2, so that they create folds that resemble numerous separate “inner walls”. In almost all instances, however, these sublayers merge to become part of the spore wall. This layer is considered a discrete structure because it appears to be homologous with a layer of similar structure and position in other species (e.g., A. koskei, A. laevis). It has been described as “semi-rigid” by Schenck and coworkers because it breaks together with the spore wall (like gw1 described below).
| Spores mounted in PVLG | |||
|---|---|---|---|
|
|
|
|

| Spores in PVLG & Melzer’s reagent | |||
|---|---|---|---|
|
|
|
|
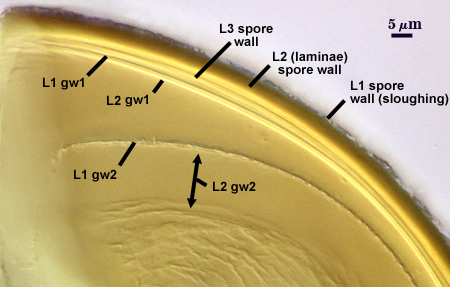
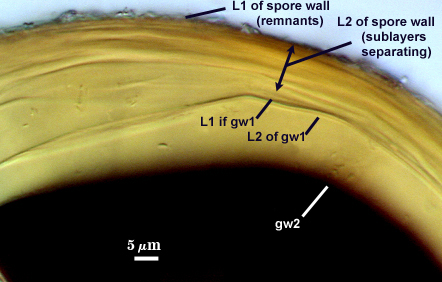
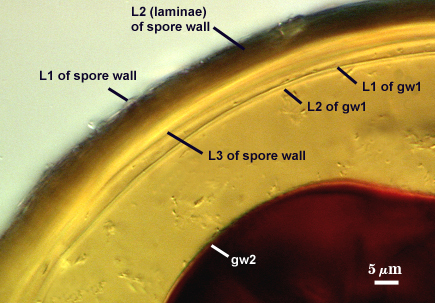
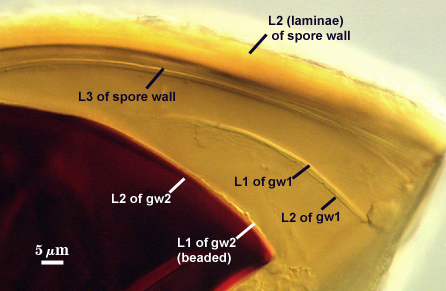
GERMINAL WALLS: Two flexible hyaline inner walls (gw1 and gw2) are present that readily separate from each other and from the spore wall.
GW1: A bilayered hyaline wall that separates easily from the spore wall and thus can be seen readily. Layers separate slightly in only a few spores, where they can be measured. L1 is less than 0.5-0.6 µm thick, folding slightly when separated from L2. L2 is < 0.5-0.7 µm, usually 0.6 µm thick. This wall has some inherent rigidity because it breaks with the spore wall (unlike iw2, which can remain intact and appear as a large “bubble”), which is why it has been described as a “semi-rigid unit wall” in the protologue.
GW2: Two adherent hyaline layers are formed. L1 is 0.6-1.2 µm thick (measurable only in PVLG), with granular excresences (or “beads”) that tend to become dislodged and float away with applied pressure. These “beads” are stabilized after preservation in formalin, but otherwise may be absent on mounted spores within a few months of storage. L2 is plastic enough that it has been termed “amorphous” (originally defined as an “amorphous wall”). It is 0.8-7.0 µm thick in PVLG-based mountants, depending on amount of pressure applied to it while breaking the spore; staining red-purple (20-80-20-0) to dark red-purple (40-80-60-0) in Melzer’s reagent.
Cicatrix
Scar indicating region of contact between spore and saccule neck during spore synthesis; very narrow lip (not measured) circumscribing the scar, circular to oval-shaped, 6.2-9.8 µm at widest (mean = 7.4 µm ).
Sporiferous Saccule
COLOR: Hyaline.
SHAPE: Mostly globose to subglobose, occasionally irregular.
SIZE DISTRIBUTION: 90-110 µm, mean = 102.8 µm
SACCULE WALL: One layer, smooth surface, 0.8-1.0 µm thick.
DISTANCE 40-80 µm.

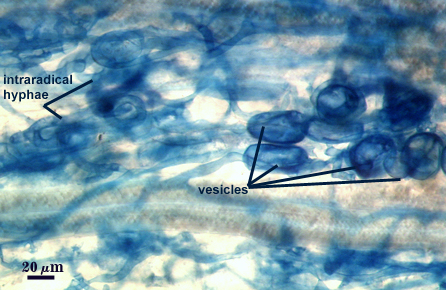
Mycorrhizae
Arbuscules and intraradical hyphae stain faintly in trypan blue, although intensity can be highly variable with age of the mycorrhizae and host plant. Infection units appear to be patchily distributed with oblong to irregular vesicles often forming in small clusters.
| Arbuscules in cortical cells of corn roots | ||
|---|---|---|
|
|
|

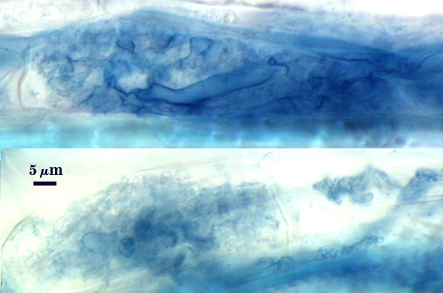
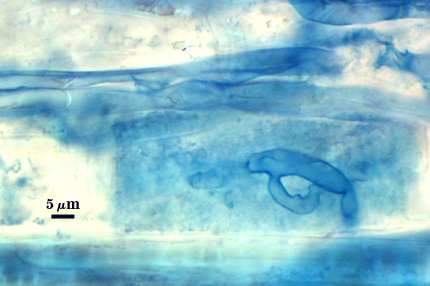
| All mycorrhizal structures in corn roots | ||
|---|---|---|
|
|
|
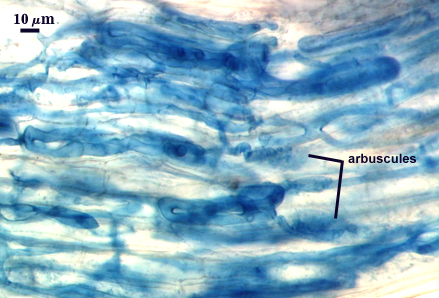
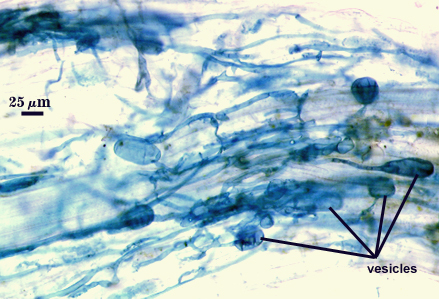
Notes
Spores rarely found with saccules attached, even in early pot culture harvests. Spore differentiation thus is thought to occur very quickly. In the description of A. mellea (Schenck et al., 1984), spores are characterized as having four inner walls: (1) a “membranous wall” closely adherent with the spore wall, synonymous with L3 of the spore wall above; (2) a “membranous wall” separable from the spore wall, synonymous with iw1 but having two adherent thin layers instead of being monolayered; (3) a beaded “membranous wall” synonymous with L1 of gw2 above; and (4) a “membranous wall” synonymous with L2 of gw2 above which was reported to produce only a light purple reaction in Melzer’s reagent. For some reason, Schenck et. al. (1984) did not correctly interpret the Melzer’s reaction of this fourth “wall” and thus failed to recognize it as being “amorphous”. Some spores in type specimens clearly produce a dark red-purple reaction to L2 of gw2, similar to that of freshly-extracted spores. The lighter reaction and reduced plasticity with applied pressure usually is observed consistently only in preserved spores that then are mounted (Morton, 1986). Unfortunately, we have no way of ascertaining the condition of the spores used to describe the species before they were mounted and characterized. It is obvious that Schenck and co-workers still were confused about the diagnostic characters of this species as late as 1990, because notes on INVAM accessions transferred from Florida indicated vacillation as to whether some isolates were A. mellea or A. morrowiae.
A reversible developmental mutant of BR983A (the only culture of INVAM which fits characteristics of type specimens of A. mellea) is subhyaline to pale straw (0-5-20-0) in color (like that of A. morrowiae). This lighter color can be attributed to a 2-3 fold decrease in spore wall thickness (0.6-2.0 µm in mature spores). When these spores are inoculated onto sorghum seedlings (either as single-spore or multi-spore inoculum), reversion to the wild-type commonly occurs. More on this mutant will be posted on this site at a later date.
Reference
- Schenck, N. C., J. L. Spain, E. Sieverding, and R. H. Howeler. 1984. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 76:685-699.
- Morton, J. B. 1986. Effects of mountants and fixatives on wall structure and Melzer’s reaction in spores of two Acaulospora species (Endogonaceae). Mycologia 78:787-794.