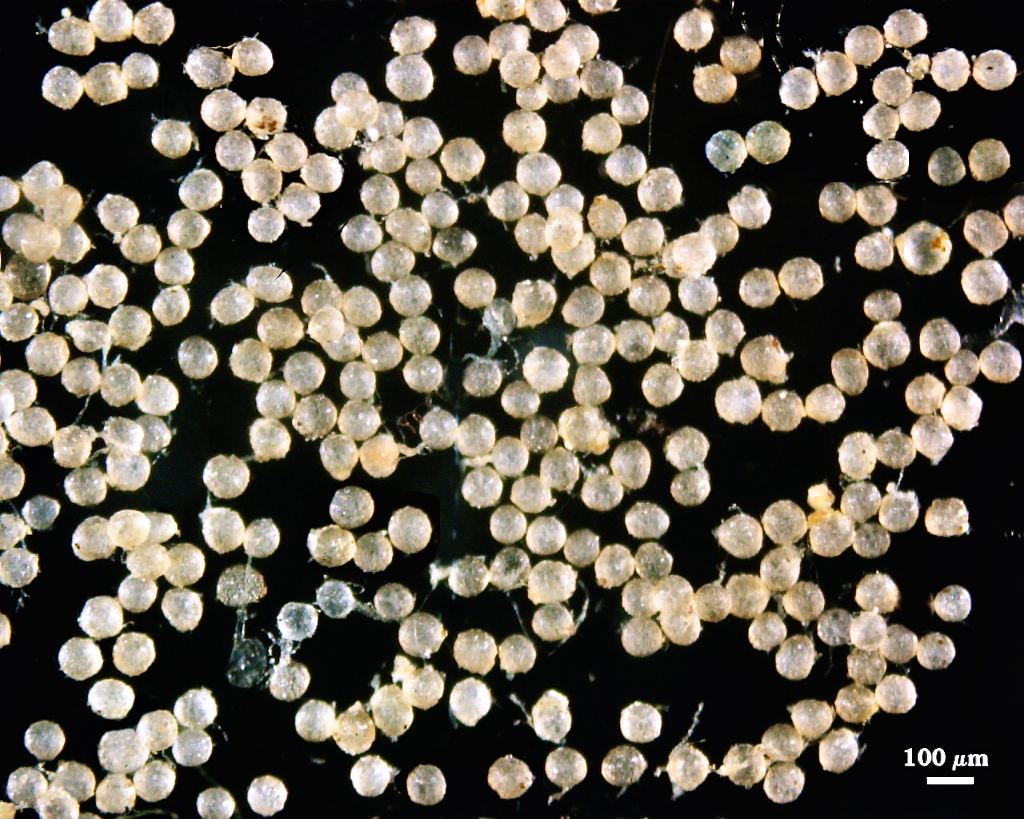
Acaulospora morrowiae
Whole Spores

 COLOR: Subhyaline (0-0-10-0) to pale yellow brown (0-15-60-0), with the former most common. Color is fairly uniform among spores in a population.
COLOR: Subhyaline (0-0-10-0) to pale yellow brown (0-15-60-0), with the former most common. Color is fairly uniform among spores in a population.
SHAPE: Mostly globose, subglobose, occasionally irregular.
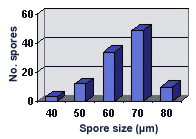
SIZE DISTRIBUTION: 60-100 µm, mean = 75.6 µm (n = 129)
Subcellular Structure of Spores
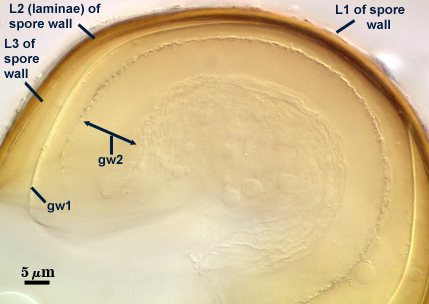
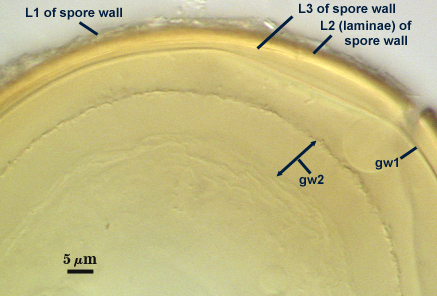
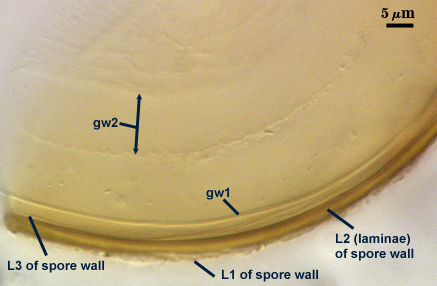
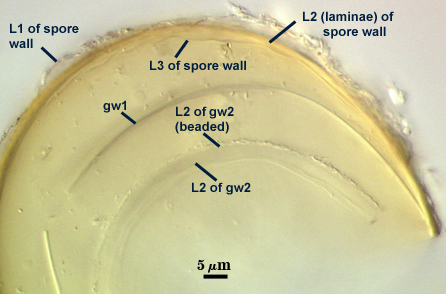
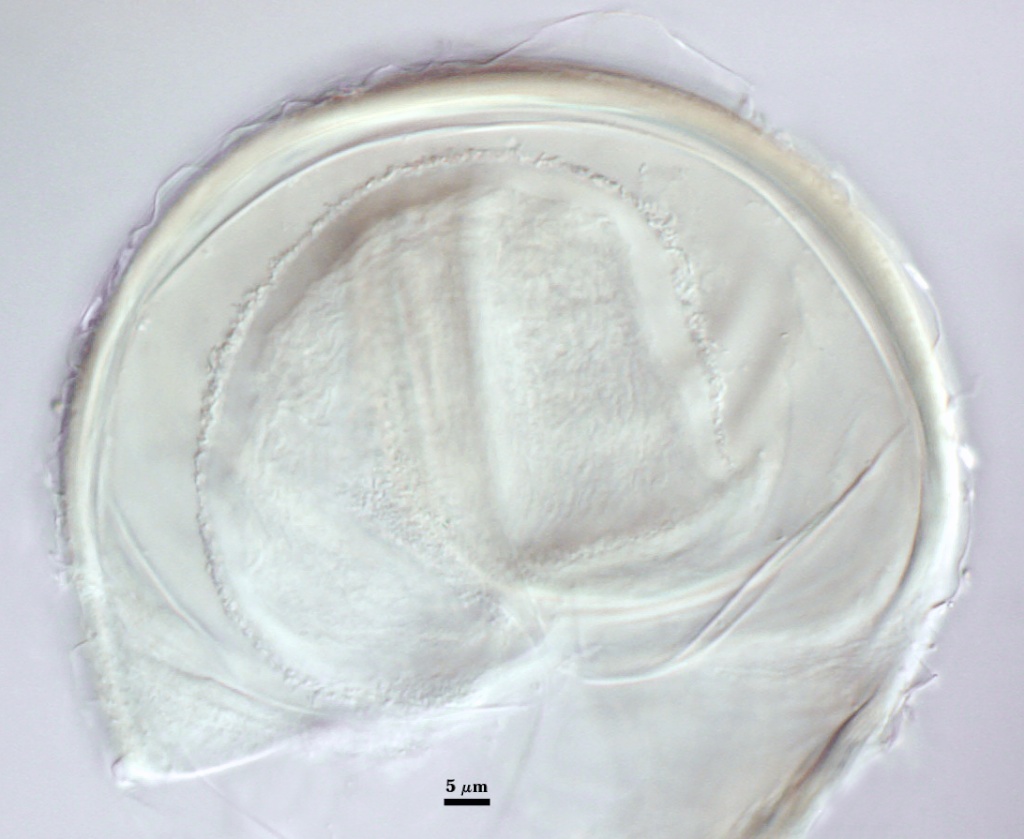
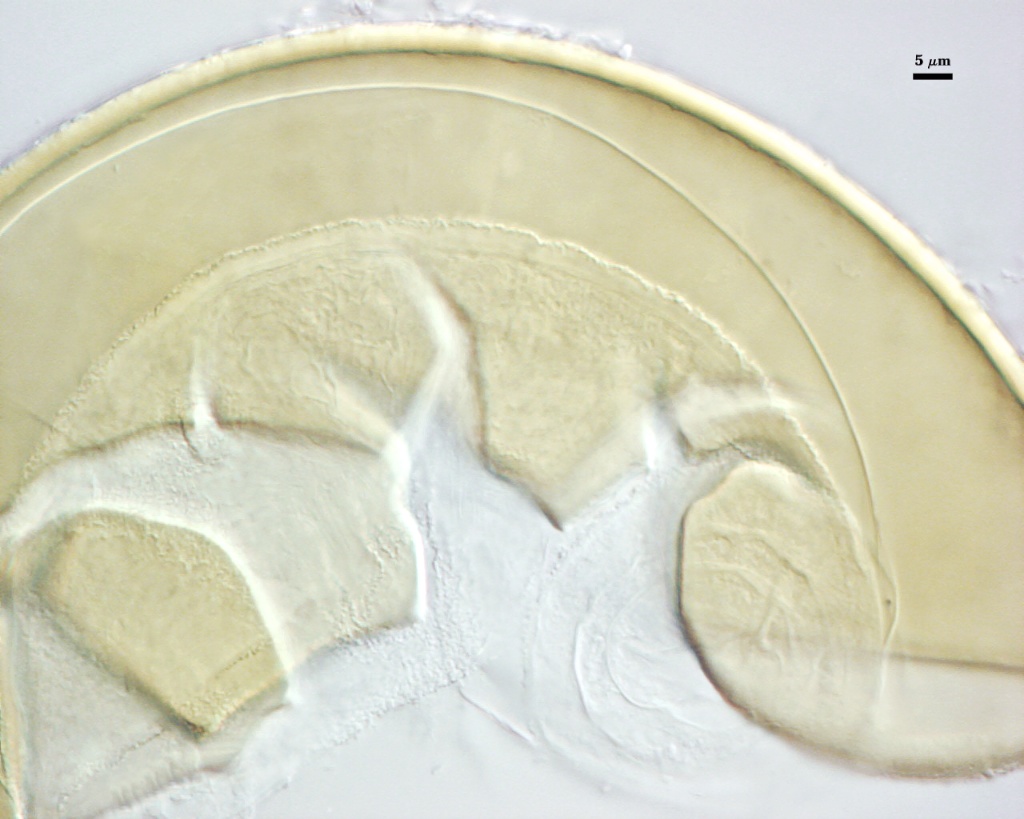
SPORE WALL: Three layers (L1, L2 and L3), the outer continous with the wall of the neck of the parent sporiferous saccule and the latter two synthesized with origination of a spore.
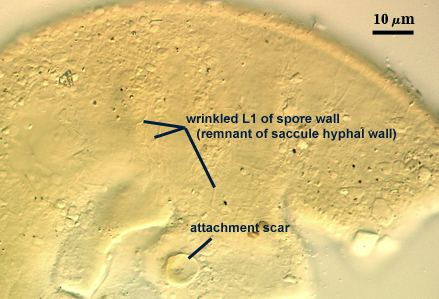
L1: Hyaline, thin (< 0.5-1 µm), and so flexible that it often produces extensive wrinkles in PVLG when present. Sloughs on many spores, particularly those collected from the field or stored pot culture inoculum. When this layer does not slough, it produces numerous folds on the spore surface and appears “rugose”. This layer has been defined as a “membranous wall”, but it clearly originates as an extension of the hyphal wall of the sporiferous saccule from which the spore wall arises and therefore is considered a component layer of the spore wall.
L2: A layer consisting of very fine and adherent sublayers (or laminae), pale yellow (0-0-20-0), 2.0-2.4 µm thick. Surface is smooth if the outer layer has sloughed, which usually occurs (especially after sucrose-density centrifugation and washing). At maturity, the pore between spore and saccule neck is closed by continuous sublayers of this layer to resemble an “endospore”.
L3: A thin layer, 0.8-1 µm thick, which often is adherent to the spore wall whereupon it remains undetected. With slight separation in some broken spores, it can be seen, but it rarely separates completely from the spore wall. We cannot determine if this layer originates with the spore wall or independent of the spore wall because of problems with limits of detection. However, we place it as part of the spore wall because a layer in the same position expands via sublayers as part of the spore wall much like behavior observed in other Acaulospora species (e.g. A. mellea, A. rehmii, and A. spinosa).
| Spores mounted in PVLG | ||
|---|---|---|
|
|
|
|
| |

| Spores in PVLG & Melzer’s reagent (1:1 v/v) | ||
|---|---|---|
|
|
|
|
| |
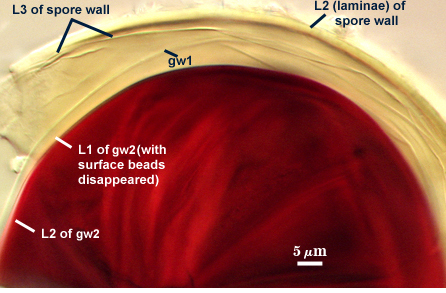
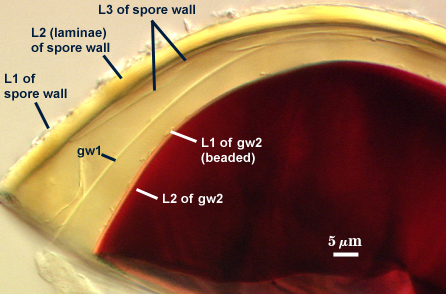
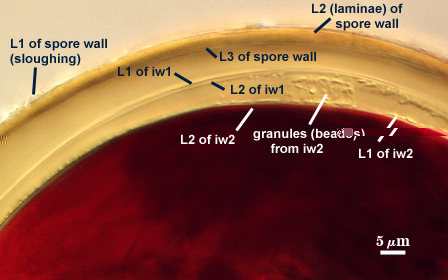
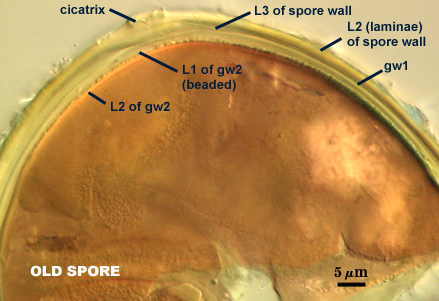
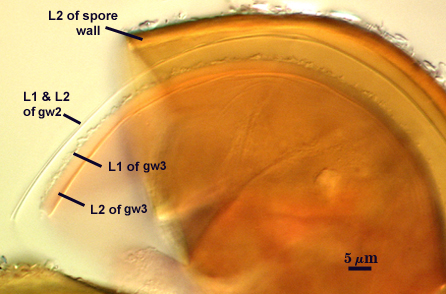
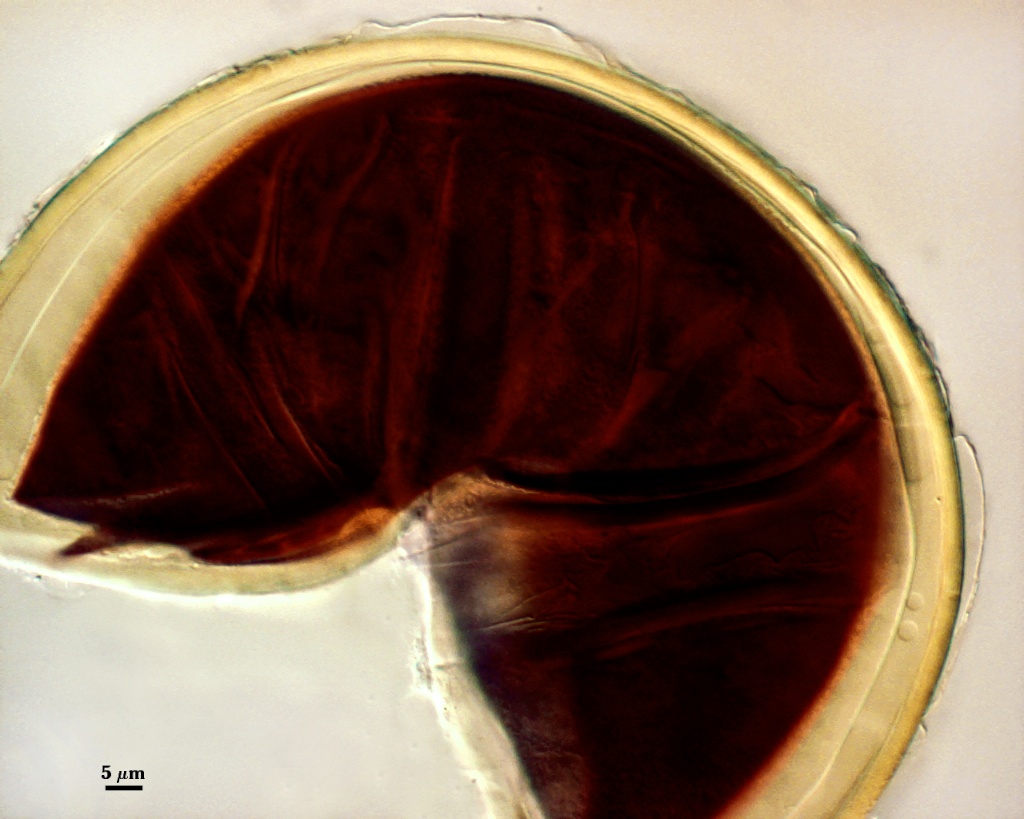
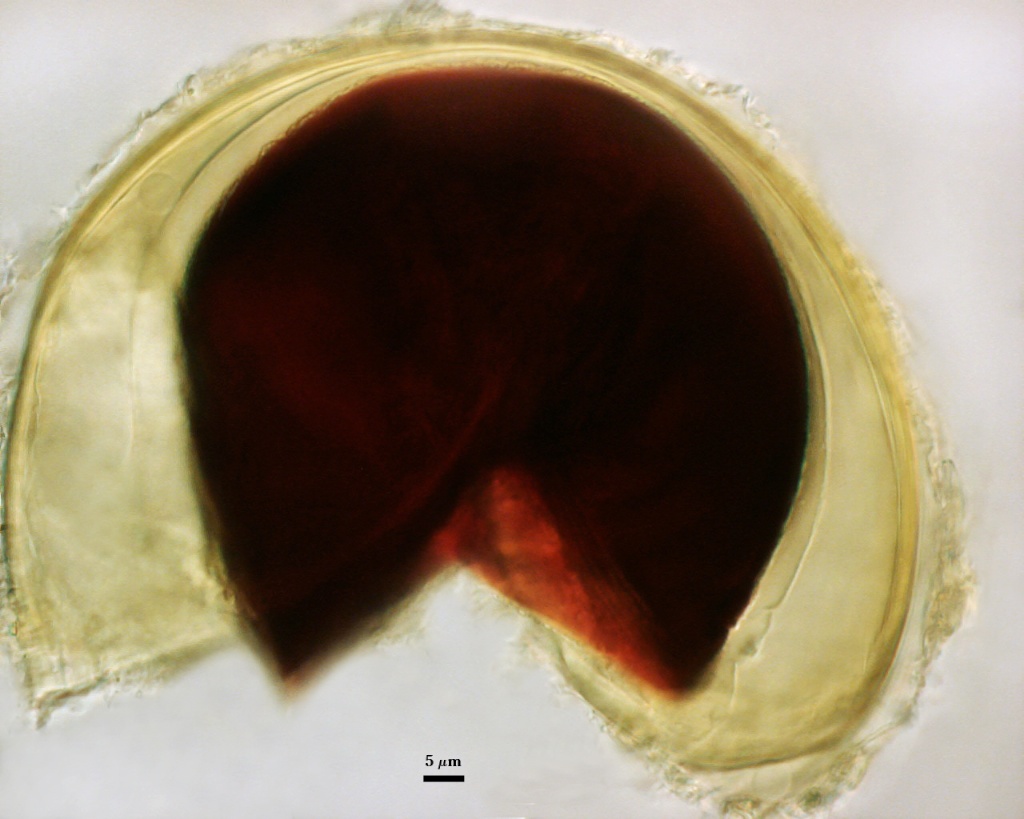
GERMINAL WALLS: Two flexible hyaline inner walls (gw1 and gw2), with gw1 sometimes obscured if it does not separate cleanly from the spore wall, but gw2 always separates and is easily identified. Folds produced by gw1 often can be deceiving and erroneously suggest additional flexible inner walls. This problem is resolved only be examining many spores, and focusing only on those in which each inner wall is broken neatly and as a single unit.
GW1: A bilayered, hyaline wall, 1-1.8 µm thick, which often separates from the spore wall when it is broken, but sometimes can remain adherent and go undetected. Both layers of this wall are of near-equal thickness, which ranges from 0.5-0.8 µm. When both layers are adherent (as they often are), this wall appears to be monolayered. However, enough spores are found in all isolates which show two distinct thin layers with differential contrast optics.
GW2: Two adherent hyaline layers are formed, which are readily distinguishable in Melzer’s reagent because of contrasting reactions. L1 is hyaline, 0.5-0.8 µm thick, with granular excresences (or “beads”) that tend to become dislodged and float away with applied pressure. These “beads” are stabilized after preservation in formalin, but otherwise may be absent on mounted spores within a few months of storage. This layer is most discretely observable when the “beads” are absent because it does not produce more than a pale pink color reaction and thus contrasts nicely from the inner L2 layer. L2 is hyaline and plastic and in conventional terminology has been described as “amorphous”. It ranges from 5-16 µm thick in PVLG-based mountants, depending on amount of pressure applied to it while breaking the spore, and stains red-purple (20-80-20-0) to dark red-purple (40-80-60-0) in Melzer’s reagent. The intensity of the reaction in Melzer’s reagent decreases considerably in old stored spores, parasitized spores, or spores preserved in lactophenol, formalin, or sodium azide (see last photo in set above).
Cicatrix
Scar indicating region of contact between spore and saccule neck during spore synthesis; very narrow lip (< 1 µm) circumscribing the scar, circular to oval-shaped, 8-10 µm at widest (mean = 8.9 µm ).
Sporiferous Saccule
COLOR: Hyaline.
SHAPE: Mostly globose to subglobose, occasionally irregular.
SIZE: 60-90 µm, mean = 72 µm.
SACCULE WALL: One layer, smooth surface, 0.8-1.0 µm thick.
DISTANCE FROM SACCULE TO SPORE: 50-110 µm.
Mycorrhizae
Arbuscules and intraradical hyphae stain with considerable variation in intensity using trypan or direct blue, even within the same infection unit of a mycorrhizal root. The photographs below are of the darker staining structures simply because they are easier to photograph and provide more detail and information. Infection units appear to be patchily distributed with oblong to irregular vesicles often forming in small clusters.
| Arbuscule in cortical cells of a corn root | ||
|---|---|---|
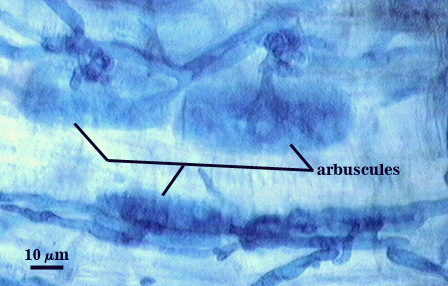 | 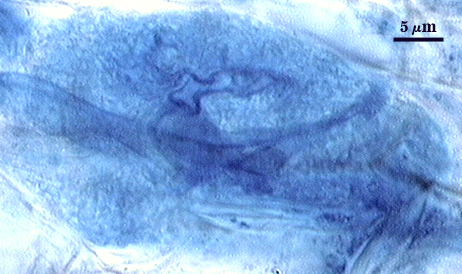 |  |
| All mycorrhizal structures in corn | |
|---|---|
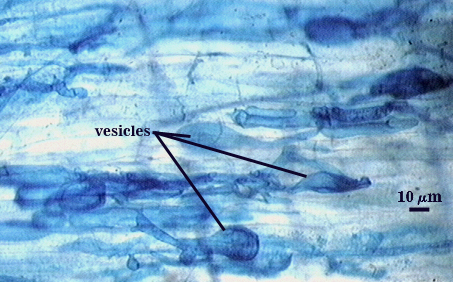 | 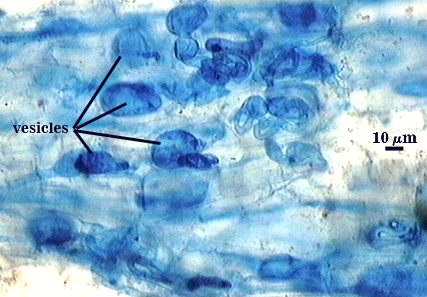 |
Germination
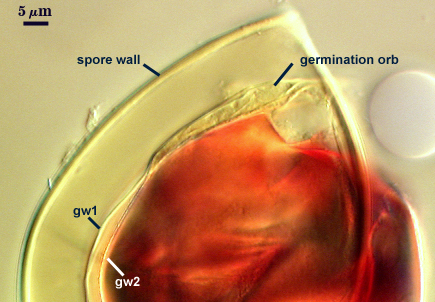
An ovoid “germination orb” forms on iw3, from which germ tubes form and penetrate through the spore wall. This orb is difficult to see except in older spores where contents have cleared with fusion of lipid globules in the spore lumen. In the photo at right, a lateral view of the orb is evident, but the margins are not resolvable. Joyce Spain has communicated that germination orbs in Acaulospora species are most visible in older spores where the contents have collapsed to some extent. We rarely see them, probably because we focus only on healthy spores.
Notes
Spores rarely found with saccules attached, even in early pot culture harvests. Spore differentiation thus is thought to occur very quickly. All available evidence indicates this species is synonymous with both Acaulospora rugosa and Acaulospora longula. The former was described as a distinct species because too much weight was given to the rugose phenotype of the outer layer of the spore wall (Morton, 1986) and the amorphous phenotype of L2 in gw2 had not been recognized or described accurately in the description of A. morrowiae. The latter was separated from A. morrowiae by length of the distance between spore and saccule and subhyaline spore color, in contrast to the bright yellow described for A. morrowiae (Schenck et al., 1984). Both of these characters overlap among all isolates of accessions inherited from Schenck’s collection, making them indistinguishable. In fact, notes on these isolates indicates Schenck either changed his diagnosis among the three for numerous isolates or specifically noted uncertainty in some of the diagnoses.
The inner layer of gw2 (L2) is described for A. morrowiae as producing a “light purple” reaction in Melzer’s reagent (Schenck et al, 1984), but mounted type specimens as well as those stored in lactophenol and then mounted in Melzer’s reagent both showed a dark red-purple reaction similar to that observed by this reference isolate. None of the voucher specimens of accessions of all three species sensu stricto deposited from the Florida collection were mounted in Melzer’s reagent, suggesting that the diagnostic value of this stain was unrecognized at the time these species were described.
Acaulospora morrowiae is a common species in many regions of North and South America and in Hawaii. Presently, 43 accessions of this species are deposited with INVAM, many from the CIAT program.
High Resolution Images | |
|---|---|
 |  |
 |  |
 |  |
 |  |
Reference
- Morton, J. B. 1986. Three new speces of Acaulospora (Endogonaceae) from high aluminum, low pH soils in West Virginia. Mycologia 78:641-648.
- Schenck, N. C., J. L. Spain, E. Sieverding, and R. H. Howeler. 1984. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 76:685-699.